Ralph Weichselbaum and colleagues explore how enhancing innate and adaptive immunity by combining radiotherapy and immune therapy could tip the balance of the host immune response to promote cure.
Radiotherapy is used in around 50–60% of cancer patients, as a curative treatment for those with localised cancer or isolated metastases, and as a palliative treatment for those with widespread disease. Data from many laboratories indicate that local radiation produces systemic, immune-mediated antitumour, and potentially antimetastatic, effects. Additionally, a combination of local radiotherapy and immune-modulation can augment local tumour control and cause distant (abscopal) antitumour effects through increased tumour-antigen release and antigen-presenting cell (APC) cross-presentation, improved dendritic cell (DC) function, and enhanced T cell priming.
Irradiation and host immune responses
Host immune status
A study using a mouse fibrosarcoma model first showed that the host immune status determines radiation-induced antitumour efficacy
(J Natl Cancer Inst 1979, 63:1229–35). A subsequent study showed a response to high-dose radiation in mouse melanoma tumours implanted into immunocompetent hosts, but not in those tumours implanted in mice lacking immune cells (Blood 2009, 114:589–95). Data also showed that radiation promoted antigen-specific T cell priming; however, paclitaxel and dacarbazine can suppress T cell priming and abolish radiation-induced tumour regression, emphasising that chemotherapy timing and composition warrants careful consideration when combined with radiotherapy. Reports from different tumour models highlight the importance of CD8+ T cell infiltration in radiotherapy effects.
Radiation enhances immune responses
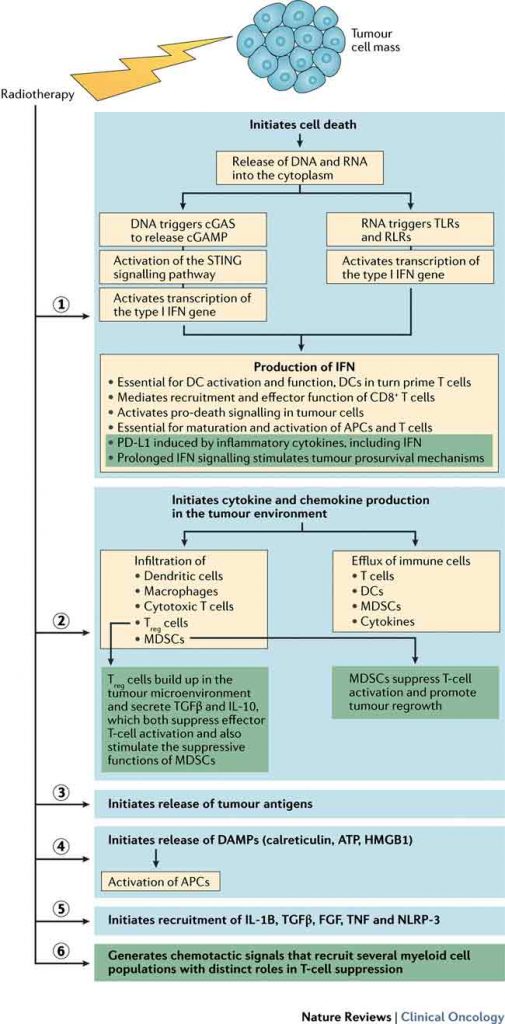
Over the past three decades, we have learnt more about how the immune system – T lymphocytes in particular – participates in tumour irradiation response. Localised radiation initiates cell death and release of cytokines and chemokines into the tumour microenvironment, leading to infiltration of DCs, macrophages, and cytotoxic T cells, and suppressor cells such as regulatory T cells (Treg) and myeloid-derived suppressor cells (MDSCs), as well as the efflux of immune cells, such as DCs, that are important antigen-presenting cells. Radiotherapy can augment innate and adaptive immune responses against tumours, decreasing immunosuppression and potentiating radiation response.
Radiotherapy can induce expression of chemokines, resulting in chemotaxis of T cells into the tumour microenvironment. Preclinical studies showed that radiotherapy-induced liberation of tumour antigens drives migration of antigen-presenting cells to lymph nodes, where T cell priming initiates a systemic response. Furthermore, localised radiotherapy induces antigen release and cross-presentation by DCs in tumour microenvironments, which can orchestrate tumour eradication following radiation, with or without immune modulation.
Research groups report that radiation changes tumour cell phenotype, resulting in upregulation of cell-surface molecules and expansion of the peptide pool, broadening antigens available for presentation, and rendering tumours more susceptible to T-cell antitumour effects. Presence of tumour-infiltrating lymphocytes (especially effector T cells) before therapy correlates with better survival in many cancers.
A study of chemoradiotherapy in rectal cancer showed that the total number of CD3+ T cells and cytotoxic CD8+ T lymphocytes is associated with disease-free and overall survival (Clin Cancer Res 2014, 20:1891–99). We speculate that radiotherapy induces release of chemokines, enriching T cell infiltrate, and enhancing T cell priming, providing positive immunological outcomes.
Natural killer (NK) cells are lymphocytes critical to host surveillance against tumours. Irradiation increases expression of NKG2D ligands in human cancer cell lines (Science 1999, 285:727–9). NK-cell-based therapies have increasingly been reported, with strategies including immune checkpoint blockade using antibodies to PD-1 or CTLA-4 and adoptive transfer of NK cells engineered to express chimeric antigen receptors (CARs) specific for tumour antigens.
In a mouse 4T1 spontaneous metastasis model, NK T-cell deficiency limited lung metastasis, and enhanced the anti-metastatic effects of radiation and anti-CTLA-4 antibody treatment (Clin Cancer Res 2009, 15:597–606).
DCs are myeloid-derived cells that are affected by alterations to tumour microenvironments from irradiation. Chemokines that attract antigen-specific T cells and DCs are released within irradiated tumours. Irradiation increases tumour-associated DCs, enhances mobilisation into draining lymph nodes, augments DC maturation, and increases the ability of DCs to cross-present antigens and prime T cells.
Activating immunosuppressive immune responses
Tumour microenvironments comprise various inhibitory immune cells (including Treg cells) and other stromal cells. Treg cells are a subset of CD4+ T cells critical for regulation of inflammation and autoimmunity, which accumulate in tumour microenvironments and secrete the cytokines TGFβ and IL-10, suppressing effector T cell activation and stimulating suppressive myeloid-derived suppressor cell (MDSC) functions. In response to localised or whole-body irradiation, Treg cell numbers increase in tumours and immune organs, which might reflect cell radioresistance. Radiation-induced increases in Treg cell numbers have been reported within tumours, but not within draining lymph nodes (Cancer Immunol Res 2015, 3:345–55). Langerhans cells (radioresistant DC subsets) can stimulate expansion of Treg cells when migrating into draining lymph nodes after whole-body irradiation (Nat Immunol 2015, 16:1060–8).
Clinical trials indicate that highly suppressive Treg cells in circulation might represent heightened immune-suppressive environments induced by chemoradiotherapy at least transiently, in patients with different cancers. Thus, targeting Treg cells, and/or the immunosuppressive effector molecule TGFβ, and CTLA-4, might reverse immunosuppression.
MDSCs contribute to tumour progression via stimulation of angiogenesis, tumour-cell invasion of adjacent tissues, and metastasis (Ann Rev Med 2015, 66:97–110). Two MDSC subsets are recognised: immature polymorphonuclear and monocytic cells.
Interactions of these inhibitory immune cell types can suppress effector T cell function and tumour angiogenesis, and promote tumour progression. MDSCs, tumour-associated macrophages, and other immune cells, are essential for tumour vascularisation.
Evidence suggests MDSCs have a role in chemoresistance and radioresistance. Of note, while MDSC recruitment is an immediate effect of radiation, a drastic reduction in MDSC numbers has been reported 7–14 days after a single high dose of radiation, and this delayed MDSC reduction could be an indirect effect of host adaptive immune response.
Tumour-associated macrophages (TAMs) are classified as immune stimulatory or immune regulatory. A mouse melanoma model showed that depletion of TAMs before irradiation increased radiotherapy antitumour effects (Cancer Res 2010, 70:1534–43), but other studies produced conflicting results, emphasising a need for additional studies into how radiotherapy affects TAMs.
Fractionation and immune cell infiltration
With fractionated radiotherapy, fraction size and timing to achieve optimal tumour effects remain to be determined. Studies combining radiotherapy with antibodies targeting a variety of immune checkpoints indicate both ablative and fractionated radiation can be effective in tumour control, depending on the experimental system studied and the approach to T cell modulation used in combination with radiation.
Role of interferon
Type-I IFN signalling in innate immune cells, such as DCs, is essential for their function to prime and activate T cells. In the setting of ablative radiotherapy, type-I IFNs improve antigen cross-presentation and T cell function.
STING signalling pathway
Stimulator of interferon genes (STING) is an endoplasmic-reticulum-associated protein that activates transcription of the type-I IFN gene. In a mouse regressing-tumour model, STING was found to be essential for a radiation-induced anti-tumour response in established tumours (Immunity 2014, 41:843–52). In vitro and in vivo studies have shown STING is required to induce type-I IFN production, and promotes antigen-specific T cell responses following radiotherapy.
When co-cultured with irradiated tumour cells, STING-deficient DCs lose the ability to produce type-I IFN and prime T cells. Ionising radiation is believed not only to kill tumour cells directly, but also to promote innate and adaptive immune responses via STING-mediated DNA-sensing pathways.
Microenvironment and tumour response
In tumour microenvironments, irradiation induces stromal, immunological and vascular changes that are essential for tumour response. Tumours have several fates: elimination, equilibrium, dormancy, and escape. Historically, relapse 5–10 years after chemoradiotherapy has been attributed to tumour dormancy, whereas long-term progression-free survival with palpable tumours after therapy cessation reflects equilibrium. Tumour dormancy involves dampening of immune processes, whereas equilibrium maintains a balanced state. Tumour escape involves evasion of immune-mediated killing mechanisms.
Tumour dormancy and equilibrium
In 1980, it was noted that presence of a palpable tumour and the rate of regression after radiotherapy did not predict cure in preclinical models or patients (Br J Cancer 1980, 4S:1–10). In immunogenic experimental models, tumour regression correlates with permanent cure; however, with non-immunogenic animal tumours, no correlation between outcome and tumour resolution at the end of treatment was observed. Hence, investigators suggested that the immune system had a key role suppressing tumours remaining palpable at the end of treatment.
Up to 50% of breast cancers and a subset of prostate cancers relapse more than five years after radiotherapy. Whether differing clinical responses reflect states of equilibrium or dormancy is unclear. One view is that radiotherapy eliminates most tumour cells, but rare radioresistant clones remain, with the extent of initial cell killing proportional to time to relapse. An alternative view is that a state of equilibrium or dormancy exists that may in part be governed by tumour angiogenesis or the host immune system.
Radiation-induced tumour equilibrium
To investigate the roles of intrinsic tumour radio sensitivity and the immune system, we studied radiation-induced tumour equilibrium and dormancy, and the contribution of adaptive and innate immunity to these processes using TUBO (HER2-positive breast cancer) and B16 (melanoma) mouse models (J Immunol 2013, 190:5874–81).
‘Early escape’ describes tumours with no response to radiotherapy; ‘stable tumours’ those that regressed and remained stable for 34–60 days; and ‘late relapse’ when formally ‘stable’ tumours regrew after 60 days. The spectrum of responses mimicked what has been observed in patients undergoing radiotherapy. The studies indicate that CD8+ T cells might mediate tumour cell death in TUBO tumours, and radiation-induced tumour equilibrium (RITE) might be a balance between cell birth and cell death. CD8+ cells are thought to mediate RITE through IFNϒ ; hence neutralisation of IFNϒ could lead to tumour regrowth. The finding that RITE exists in stable mouse tumours suggests that therapy-induced equilibrium between cancer-cell division and host immune system killing dictates disease status, supporting the feasibility of treating stable disease by tipping the balance (equilibrium) in favour of host antitumour immunity.
Data from tumour models indicates PD-L1 expression is inducible in tumour cells or host immune cells. When PD-L1 was blocked in mice harbouring stable tumours, most tumours regressed, confirming that activation of antitumour immunity can shift the balance towards eradication of non-progressing tumours (J Immunol 2013, 190:5874–81).
Notably, in a clinical trial involving 277 patients with a range of cancer types, PD-L1 expression on both tumour and immune cells strongly predicted response to PD-L1 inhibitors (Nature 2014, 515:563–7). In a mouse model, radiotherapy and same-day PD-1/PD-L1 inhibition achieved better tumour response than delayed administration (Cancer Res 2014, 74:5458–68).
Although clinical confirmation is required, we hypothesise the RITE model is applicable to some patients with radiotherapy-induced stable tumours that do not progress, who might achieve tumour resolution with immune-checkpoint blockade or immune-modulators before relapse. Mouse models are, however, heterogeneous, and not all mice developed PD-L1 expression leading to RITE.
The major clinical successes in radio-immunomodulation are the result of the advent of immune checkpoint inhibitors. It has been noted that the PD-1/PD-L1 pathway primarily regulates ongoing inflammatory activity, whereas the CTLA-4 pathway regulates autoreactive T cell responses. Patients with ‘good immune scores’ and pre-existing tumour-specific T cells have better outcomes following immune checkpoint therapy (Nature 2014, 515:496–8), and in patients who would otherwise not respond to immune checkpoint inhibitors, localised radiotherapy can induce tumour-specific T cells, promoting responses (Vaccine 2015, 33:7415–22).
Inhibition of CTLA-4 checkpoint
Inhibitors of the CTLA-4 pathway (e.g. ipilimumab) have shown encouraging results in cancer. CTLA-4 functions as an immune suppressor by increasing the signal intensity required for CD8+ T cells to engage target tumour cells.
Treatment of mouse 4T1 primary mammary carcinomas with irradiation and CTLA-4 blockade inhibited lung metastases, with therapeutic effects requiring CD8+, but not CD4+, T cells (Clin Cancer Res 2005, 11:728–34).
Results from a mouse model indicated a hypofractionated regimen plus anti-CTLA-4 therapy was more effective than either alone in inducing immune infiltrate and abscopal effects (Clin Cancer Res 2009, 15:5379–88). Results of early clinical studies of anti-CTLA-4 antibodies demonstrate improved overall survival (OS) in a range of cancers. Anti-CTLA-4 therapies have been tested in combination with other treatments, including vaccines, granulocyte-macrophage colony-stimulating factor (GMCSF), other checkpoint inhibitors, radiotherapy and chemotherapy.
A phase III trial to assess radiotherapy plus ipilimumab or placebo in patients with metastatic castration-resistant prostate cancer found no significant difference in OS, but a post hoc subgroup analysis revealed a trend towards improved OS, and significance in patients with bone metastasis (Lancet Oncol 2014, 15:700–12). In preclinical models, combined radiotherapy and anti-CTLA-4 treatment was efficacious owing to stress-induced NKG2D (natural-killer group 2, member D) expression on tumour cells that survive irradiation, making them susceptible to NK-cell-mediated cytotoxicity (Oncoimmunology 2013, 2:e23127).
Inhibition of the PD-1/PD-L1 checkpoint
PD-L1 expression has been observed in solid malignancies, and might be a dominant mechanism of immunosuppression in some tumours.
The presence of PD-L1 in tumours predicts responses to PD-1/PD-L1 blockade, and inhibition of the PD-1/PD-L1 pathway on T cells is associated with antitumour activity in mouse models and clinical trials. A mouse model showed substantial tumour regression with high-dose radiation and an anti-PD-L1 antibody. Dramatic reductions of MDSCs were seen, associated with increased CD8+ T cell infiltration and priming (J Clin Invest 2014, 124:687–95). Conversely, when CD8+ cells were depleted, MDSC recovered. MDSC-mediated suppression of T cell function in cancer progression is well-established, and MDSCs are associated with chemo-resistance.
In vitro cytotoxicity assays revealed that activated CD8+ T cells kill MDSCs directly, with cytokines produced by activated T cells leading to MDSC apoptosis. By contrast, although IFNϒ production correlates with T cell activity, IFNϒ did not mediate induction of MDSC death in assays.These reports suggest that, in preclinical models, inhibition of the PD-1/PD-L1 checkpoint combined with radiotherapy liberates T cells from immunosuppression, which in turn positively alters the tumour microenvironment.
Several mouse solid tumour models demonstrate synergistic effects of radiotherapy and immunotherapy via checkpoint inhibition, including improved survival using PD-1 inhibitors plus stereotactic radiation in orthotopic brain tumours, compared to either treatment alone, and improved survival adding PD-1 inhibitors to TGFβ blockade plus radiation-induced vaccination, compared with radiation-induced vaccination plus TGFβ blockade alone.
Radiation-induced vaccination invol–ves conversion of tumour into an in situ vaccine by inducing immunogenic death of cancer cells, promoting a pro-immunogenic tumour microenvironment and thereby priming tumour-specific T cells (Int J Radiat Oncol Biol Phys 2012, 84:879–80).
Data indicate radiation and anti-PD-1 therapy increases memory CD8+ T cell numbers. Although NK cells might contribute to local tumour control, CD8+ T cells have been shown to be required for the antitumour effects produced by combined radiotherapy and anti-PD-L1 antibodies (Cancer Res 2014, 74:5458–68).
A preliminary preclinical report indicated that radiotherapy combined with an anti-PD-1 antibody can result in primary tumour control and abscopal effects (Int J Radiat Biol Phys 2014, 90:S1), with further studies indicating that the combination induces endogenous antigen-specific immune responses, resulting in improved local control in melanoma and breast carcinoma models (Oncol 2015, 29:331–40). Timing of anti-PD-L1 blockade is crucial: concurrent but not sequential radiation results in long-term tumour control (Oncoimmunology 2015, 4:e1016709). Currently, over a dozen clinical trials are evaluating anti-PD-1 and anti-PD-L1 antibodies combined with radiation.
The abscopal effect
The abscopal effect (regression or disappearance of lesions outside irradiated fields) is rare with radiotherapy alone, but is increasingly reported when radiation is administered with immune modifiers in preclinical models and patients, leading to suggestions that it is an immune-driven phenomenon, indicating that local radiotherapy produces systemic effects. Studies show localised radiotherapy plus CTLA-4 blockade inhibited development of lung metastases; and radiotherapy plus anti-PD1 antibody treatment produced abscopal effects in a range of carcinoma models, with one study showing almost complete regression of the primary tumours and a 66% reduction in distant tumours.
Several case reports highlight regression of targeted lesions and abscopal effects in melanoma treated with ipilimumab and radiotherapy (NEJM 2012, 366:925–31). In the first clinical trial testing abscopal responses with radiotherapy and GMCSF, 27% of patients with metastatic solid tumours experienced the desired effect (Lancet Oncol 2015, 16:795–803). Collectively, observations of the molecular and cellular events generating the abscopal effect indicate these are the result of a cellular feedback mechanism involving effector T cells within the irradiated tumour microenvironment, which occurs several days after local radiotherapy and subsequent to APC migration and T cell activation in the draining lymph nodes. Thus, the abscopal effects could be modulated by tipping the balance between positive immune regulators (of T cell function) and negative regulators (of the local and systemic suppressive microenvironment) to elicit strong immune responses.
Conclusions
Current evidence indicates that radiotherapy can invoke both local and systemic immune responses, which can either support tumour cell survival or promote tumour cell death. Enhancing innate and adaptive immunity by combining radiotherapy and immunotherapy is thus crucial to improve patient survival. Current radioimmunotherapy paradigms are based on results from animal models, observations of responses in patients, and preliminary data from trials of combination immunotherapy and radiotherapy. Robust hypothesis-testing clinical trials are required to determine the appropriate approach to integrating these modalities.
Key points
Radiotherapy not only exerts direct cytotoxic effects on tumour cells, but also reprogrammes the tumour microenvironment to exert a potent antitumour immune response.
Tumour cell proliferation and cell death due to T cell cytotoxic killing coexist in irradiated tumours, resulting in stable disease that might provide a window of opportunity for immune modulation.
Radiotherapy enhances antitumour immunity, but also induces immunosuppressive responses.
The combination of immunotherapy and radiotherapy presents a multimodal treatment approach that involves stimulating and suppressing various pathways.



This is a summary, by Janet Fricker, of a longer article that was first published in Nature Reviews Clinical Oncology: Ralph R. Weichselbaum, Hua Liang, Liufu Deng & Yang-Xin Fu (2017) Radiotherapy and immunotherapy: a beneficial liaison? Nat Rev Clin Oncol vol. 14, pp 365–379, doi:10.1038/nrclinonc.2016.211, and has been modified with permission. © Macmillan Publishers Ltd