Sriram Venneti and colleagues examine non-invasive metabolic imaging strategies that can be used to interrogate some of the genomic alterations in brain tumours, with the ultimate goal of informing patient management.
Clinical management of brain cancer relies heavily on non-invasive imaging because location often prohibits tumour sampling and/or surgical resection. MRI (with or without contrast enhancement) remains the standard approach and is used to guide tissue biopsy sampling, establish diagnosis, assess progression, and evaluate therapy effectiveness.
Work is needed to exploit knowledge of brain tumour pathology and biology to develop non-invasive imaging modalities guiding diagnosis, treatment and follow-up monitoring of brain cancer.
In the personalised medicine era, we are beginning to appreciate the genetic complexity of primary brain tumours and uncover a variety of clinically relevant mutations. Such information allows identification of actionable targets and development of corresponding molecularly targeted therapies. Challenges remain in identifying, stratifying, and monitoring patients using advanced imaging technologies, and leveraging information gathered from genomics.
Cancer metabolism provides the best example of molecular imaging in cancer patients, with PET imaging and magnetic resonance spectroscopic imaging (MRS) being the principal modalities used.
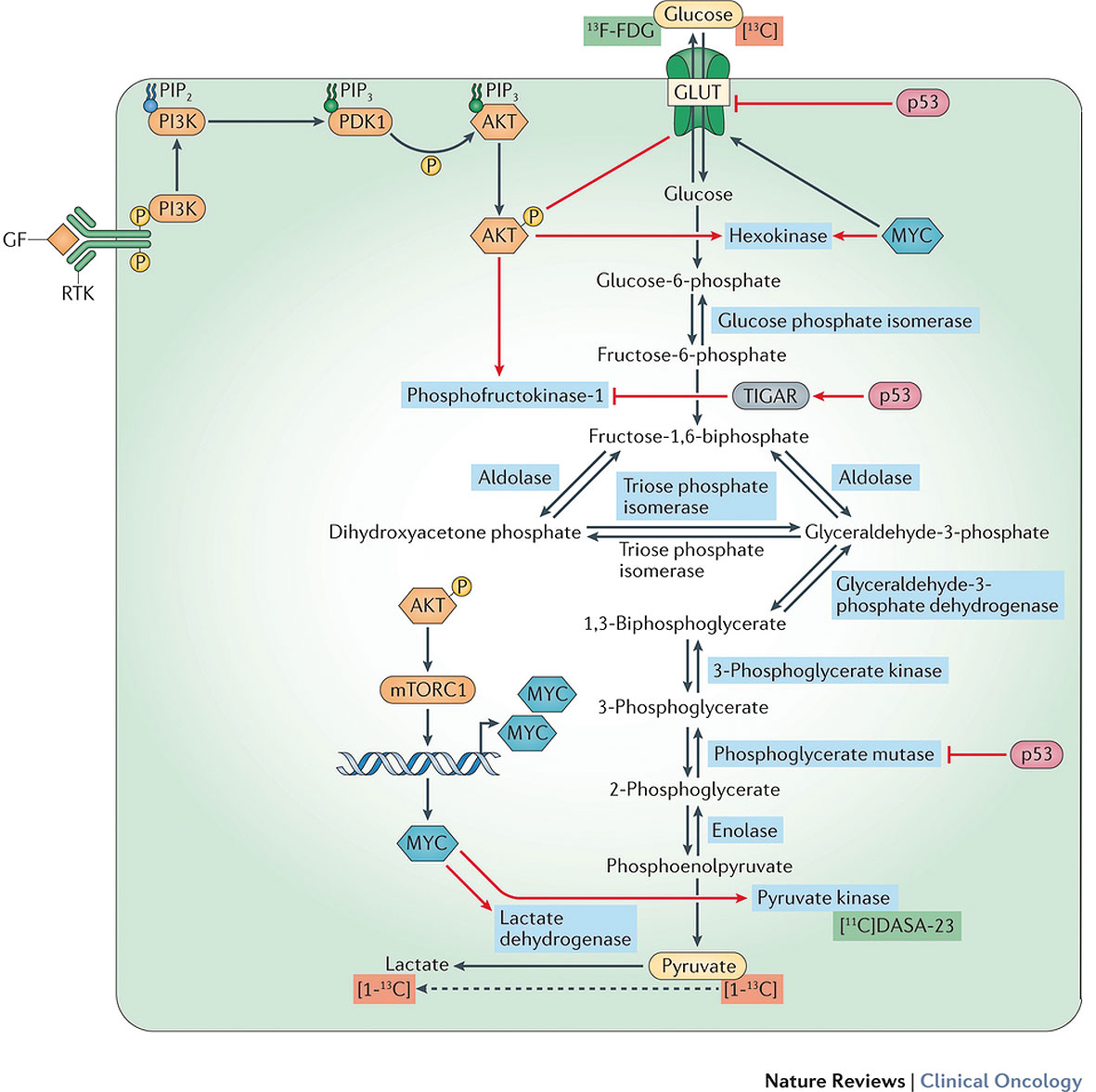
PET imaging, based on biological substrates labelled with radionuclides, is highly sensitive, with signal detection limits of 10-11 to 10-12 mol/l (Curr Pharm Biotechnol 2010, 11:555–71). It uses the ‘Warburg effect’, where cancer cells generate ATP predominantly through aerobic glycolysis of glucose to lactate in the cytoplasm. The number of ATP molecules produced (2 molecules) is lower than mitochondrial metabolism (36 molecules), increasing glucose demands of cancer cells. PET imaging uses an 18F-labelled glucose-analogue tracer (18F-FDG) that cannot be metabolised further than initial phosphorylation by hexokinase, and therefore accumulates in cells, indicating glucose demand.
Single-voxel MRS and multi-voxel MRS (MRSI) characterise the chemical and molecular composition of tumours based on radiofrequency signals generated by nuclear spins of magnetic-resonance-active nuclei, including 1H, 31P, and 13C (Semin Oncol 2011, 38:26–41).
The capacity of oncogenes to reprogramme cellular metabolism (enabling tumour cells to survive, grow and proliferate) is emerging as a fundamental concept in cancer biology. Many oncogenes expressed in brain tumours influence specific metabolic pathways, including glucose, amino acid, and fatty metabolism.
Genomic heterogeneity of glial tumours
The updated 2016 WHO Classification of Tumours of the Central Nervous System defines two major glial tumour groups: astrocytic tumours and oligodendroglial tumours (IARC, 2016). Observations from next generation sequencing studies that adult and paediatric glial tumours are genetically distinct prompted WHO to incorporate molecular characteristics, histological type and tumour grade into the revised classification system.
For example, in adults >90% of glioblastoma multiforme (GBM) harbour genetic alterations converging on the P13K/AKT/mTOR pathway, including enhanced activation of receptor tyrosine kinases transmitting signals via this pathway (such as EGFR, PDGFRA, and MET). By contrast, high-grade gliomas in children are characterised by mutations resulting in epigenetic reprogramming, with histone H3 K27M and G34R/V mutations detected in ~60% of paediatric patients with GBMs. Thus, genetic drivers of childhood and adult gliomas are distinctly different.
Precision medicine for brain tumours
Subclassification of brain tumours into distinct groups informing management of patients beyond histological grade is another important development arising from molecular tumour studies. For example, grade IV medulloblastomas (predominantly paediatric) are classified into four molecular subtypes: Sonic Hedgehog, WNT, Group 3 and Group 4.
Next generation sequencing studies have refined understanding of brain tumours and patient care, with research opening novel avenues for targeted therapies. For example, vismodegib (an antagonist of the SHH pathway) is effective in patients with SHH medulloblastoma, but not other subtypes (JCO 2015, 33:2646–54).
Integrating molecular characteristics of brain tumours into therapeutic decisions is increasingly integral to patient management, with molecular features of brain cancers currently assessed using tissue samples obtained from tumour biopsy or resection. Obtaining such samples is not always feasible for pontine and brainstem tumours, owing to surgical challenges involving location. Furthermore, longitudinal assessment of serial samples is not practical due to difficulties performing repeated biopsies.
Since conventional MRI is inadequate for evaluating molecular alterations, non-invasive, metabolic imaging of brain tumours is emerging as a way to assess molecular and metabolic alterations.
Imaging glucose metabolism
Glucose is metabolised into pyruvate by glycolysis, which under aerobic conditions can be oxidised to acetyl-coenzyme A, which then enters the mitochondrial tricarboxylic acid cycle, fuelling ATP production via oxidative phosphorylation. Brain tumours typically exhibit the Warburg effect, where pyruvate is diverted from the mitochondria and converted to lactate (NMR Biomed 2012, 25:1234–44). Enhanced production of glycolytic intermediaries promotes tumour invasion and escape from immune cells, and glycolytic metabolism enables adaptation to low oxygen levels in hypoxic areas of the tumour.
In brain tumours, the Warburg effect is controlled at several levels, including regulation by the P13K/ AKT/ mTOR pathway via BRAF activation.
Interestingly, many glycolytic enzyme isoforms expressed in brain development are expressed in brain tumours, but not the adult brain, e.g. HK2 and pyruvate kinase M2 (PKM2). Such observations have enabled the development of a 11C-labelled PET imaging probe specific for PKM2 (Clin Cancer Res 2005, 11:2785–808).
18F-FDG-PET imaging was one of the earliest tools measuring locations of brain glucose utilisation. However, in gliomas 18F-FDG-PET has poor tumour-to-background contrast, due to high levels of glucose uptake in normal brains. Using 13C-labelled glucose as a tracer (quantifying the appearance of 13C-glucose in lactate or pyruvate), MRS can monitor glucose metabolism in glioma.
To overcome low signal-to-noise ratios, 13C-enriched probes can be ‘hyperpolarised’ through exposure to low temperature microwaves, altering the Boltzmann distribution of 13C, and increasing MRS detection more than 10,000 fold (Proc Natl Acad Sci 2003, 100:10158–63). In small animal models, generation of [1-13C] lactate from [1-13C] pyruvate was detected in glioma xenografts in vivo, but not in non-tumour brain tissue.
The spectrum of metabolic imaging studies might be further expanded by spectroscopic techniques with enhanced physiological and functional information based on CEST contrast, a sensitivity enhancement mechanism by which low concentrations of solutes containing exchangeable protons that have different resonance frequencies from bulk water protons can be selectively saturated with radiofrequency energy and visualised indirectly using the water signal (Magn Reson Med 2011, 65:927–48).
Finally, combined PET–MRI systems are enabling a multiparametric approach to non-invasive brain tumour characterisation, although further studies are needed to optimise this approach in the clinical setting.
Oncogenic reprogramming and imaging of glycolysis in brain tumours
Red arrows depict common oncogenic signalling pathways in brain tumours, such as those involving activation of PI3K, AKT, mTOR and MYC, and those influenced by inactivating alterations in p53, which affect various aspects of glycolysis — generally increasing the glycolytic flux. PET tracers relating to this pathway are highlighted in green, and magnetic resonance spectroscopy (MRS) tracers in red. The two main substrates involved in glycolysis and related pathways that can be labelled for use as clinical imaging tracers are glucose and pyruvate (yellow ovals). [click to enlarge]
Imaging of IDH-mutant gliomas
Human cells express three isoforms of IDH (enzymes converting isocitrate to α-keloglutarate): IDH1, IDH2, and IDH3. IDH1 is present in the cytosol and IDH2 and IDH3 in the mitochondria. Since the metabolite D-2-hydroxy-glutarate (D-2HG) is produced in IDH1/2 mutant gliomas, but not wild-type tumours, the metabolite could serve as a biomarker for diagnosis, treatment and surveillance via non-invasive imaging.
The detection of D-2HF in vivo using MRS correlates with better prognosis compared with patients in whom it is not detected. Sensitivity, however, depends on tumour size – 8% for small tumours versus 91% for larger tumours (Neuro Oncol 2016, 18:283–90). In addition, hyperpolarised-13C MRS might be harnessed to probe for the enzymatic function of mutant IDH1/2 in vivo (Nat Commun 2013, 4:2429).
Imaging amino acid metabolism
Mechanisms where non-invasive imaging of brain tumours could exploit reprogramming of amino acid metabolism include:
Glutamate: Many primary brain tumours (including gliomas, meningiomas, and medulloblastomas) have altered levels of glutamate and glutamine. Tumour cells also exchange glutamate for cysteine (via the cysteine-glutamate anti-porter), raising the possibility that PET imaging of glutamate exchange could analyse brain tumours.
Glutamine: Brain tumours display increased cellular glutamine uptake and metabolism. Non-invasive in vivo measurement of glutamine uptake can be achieved using 18F-FGln, which has been shown to increase in gliomas compared to normal brain tissue (Sci Transl Med 2015, 7:274ra)
Methionine: 11C methionine uptake is increased in malignant cells, including gliomas. It can be useful in detecting tumours, assessing treatment response and predicting disease recurrence.
Aspartate: Levels of NAA (synthesised in neuronal mitochondria from aspartate and acetyl-CoA) decrease in gliomas, most probably due to reduced expression of enzymes involved in NAA biosynthesis. 1H-MRS can be used to detect decreases in NAA.
Imaging fatty acid metabolism
Cancer cells undertake both de novo fatty acid synthesis and enhanced fatty acid oxidation (Nat Rev Clin Oncol 2017, 14:11–31). Choline is an essential nutrient required for synthesis of phospholipids and the neurotransmitter acetylcholine, and levels of this metabolite are often increased in cancer cells, which might reflect increased rates of cell membrane turnover (Semin Oncol 2011, 38:26–41). Elevated total choline signals can be used clinically for primary diagnosis of low-grade and high-grade glioma, and detection of recurrent glioma versus tumour necrosis following radiation therapy.
Molecular imaging of other processes
Cell proliferation: 18F-FLT-PET might be used to detect highly proliferative tumours, reflecting dependence on de novo thymidine synthesis and the thymidine salvage pathway.
Hypoxia: Hypoxia is a key feature of rapidly growing tumours that promotes adaptive responses (such as angiogenesis). 18F-FMISO-PET enables spatial delineation of hypoxic regions of brain tumours (J Nucl Med 2004, 45:1851–59).
Angiogenesis: Angiogenesis is another hallmark of tumour growth. αVβ3-integrin, expressed on endothelial cells during angiogenesis, offers a target for PET scanning using ligands with high affinities for αVβ3-integrin (Eur J Nucl Med Mol Imaging 2010, 37:S86–103).
Metabolic imaging in patient care
In the following section, the potential for metabolic imaging to be assimilated into routine clinical care is explored.
Tumour delineation for local therapy: Use of amino-acid based PET can improve identification of most biologically aggressive components of heterogeneous low-grade and high-grade gliomas – information that could dictate subsequent therapy and reduce incomplete resection. Use of multiple imaging modalities, including multi-voxel 1H-MRS and diffusion and/or perfusion MRI, can also help distinguish between heterogeneous regions of dense tumour infiltration, areas of oedema with admixed tumour cells, and oedema not infiltrated by tumour (Neuroradiology 2006, 48: 622–31).
Prognostication and response prediction: In patients with low-grade glioma, 18F-FET-PET time-activity curves correlate with malignant progression and survival, suggesting a prognostication role. Use of H-MRS for predicting survival outcomes has been evaluated in multiple studies in adult and paediatric patients with brain tumours, with worse outcomes observed in patients with an elevated tumour choline-to-NAA ratio before treatment or adjuvant chemotherapy.
Assessing treatment response: A variety of metabolic changes in both the tumour and microenvironment are observed following cytotoxic, radiation and antiangiogenic therapies. MRS imaging of adjuvant radiation in malignant gliomas, for example, reveals declines in mean tumour choline-to-NAA ratio that predict outcome (Clin Invest Med 2006, 29:201–11) Such findings open opportunities for adaptive, response-based radiation treatment.
Monitoring of resistance or progression: Both glucose and amino acid metabolism undergo reprogramming during emergence of resistance to radiation and chemotherapy. Thus, non-invasive metabolic imaging has potential to detect treatment resistance at an early stage and inform alterations in clinical care.
Molecular subgrouping: The non-invasive identification of clinically relevant (prognostic or predictive) molecular tumour subgroups offers a potential application for metabolic imaging. Use of MRS, for example, allows differentiation between SHH and Group 3/4 medulloblastomas.
Conclusions
Altered cancer metabolism offers a unique window to integrate genomic information with advanced imaging modalities. The field has progressed, with novel techniques implemented for analysis of tumours in preclinical models and patients.
Additional work is needed to standardise these metabolic imaging techniques for routine clinical use. In combination with other functional imaging modalities, these techniques might prove complementary to conventional MRI in characterising tumour biology and metabolism, with the aim of informing patient management. Future studies based on oncogene-driven metabolic pathways, might enhance diagnosis, prognostication, treatment and surveillance of brain tumours, ultimately improving patient outcomes.
Take home messages from the authors
From the left: Michelle M Kim is affiliated to the Department of Radiation Oncology and Abhijit Parolia and Sriram Venneti to the Department of Pathology, of the University of Michigan Health System, University of Michigan, Ann Arbor, Michigan; Mark P. Dunphy is affiliated to the Molecular Imaging and Therapy Service of the Department of Radiology, at the Memorial Sloan Kettering Cancer Center, New York.
“We have made significant progress in understanding the genetics of brain tumours. However, we have not devised ways to detect these tumours non-invasively in living patients. These genetic alterations in brain tumours can reprogramme metabolic pathways in cells. Our aim was to examine if metabolic imaging could help better understand and evaluate brain tumours in the clinic.
Our main take home messages are two-fold. First, there is great promise in metabolic imaging to detect molecular alterations. For example altered metabolism in isocitrate dehydrogenase 1/2 mutant gliomas can be detected using a technique called magnetic resonance spectroscopic imaging (MRS). Second, a lot of research will need to be done to come up with such techniques for other tumour mutations and to standardise these imaging techniques for daily clinical practice.
Clinical implications
Metabolic imaging could help with non-invasive diagnosis, monitoring and assessment of the therapeutic efficacies of treatment. This would be of enormous benefit for patients.
Further studies
The field of metabolic imaging holds great potential. We need to develop more novel techniques, and many of the existing imaging techniques, such as hyperpolarised MRS, need to be considered more carefully in order to directly impact patient care.




This is an abridged version of Michelle M. Kim et al. (2016) Non-invasive metabolic imaging of brain tumours in the era of precision medicine. Nat Rev Clin Oncol 13, 725–739, doi:10.1038/nrclinonc.2016.108. It was edited by Janet Fricker and is published with permission © Macmillan Publishers Ltd.